低频突变能够提供重要的诊断或治疗信息,包括癌症、产前诊断、传染病和器官移植等领域。由于低频突变的丰度低,常常会被大量存在的wtDNA(wild-type DNA)淹没,较难检测,而wtDNA提供的信息又非常有限。
因此,自20世纪90年代初以来,低频突变特异性捕获检测技术(简称mutEnrich技术)开始发展,特别是随着用于癌症残留病灶监测、治疗耐药性检测以及早期癌症检测的液体活检相关研究的深入,迫切需要实用、有效、能够用于检测临床相关低频突变的mutEnrich技术,随着方法学优化和新技术的出现,mutEnrich技术的检测性能也正在得到持续的改进和优化。
2022年10月,《Clinical Chemistry》发表mutEnrich技术的综述文章,来自哈佛医学院和Broad研究所的科学家们回顾了2010年以来的mutEnrich技术[1],并将mutEnrich技术分三个类别(PCR Blocker、Enzymatic、Physicochemical以及与NGS技术的结合)进行了系统总结。本文结合综述文章,对Enzymatic技术部分进行介绍。在过去的十多年中,具有更广泛催化能力的核酸内切酶提供了强大的低频突变富集和检测工具(表1)。
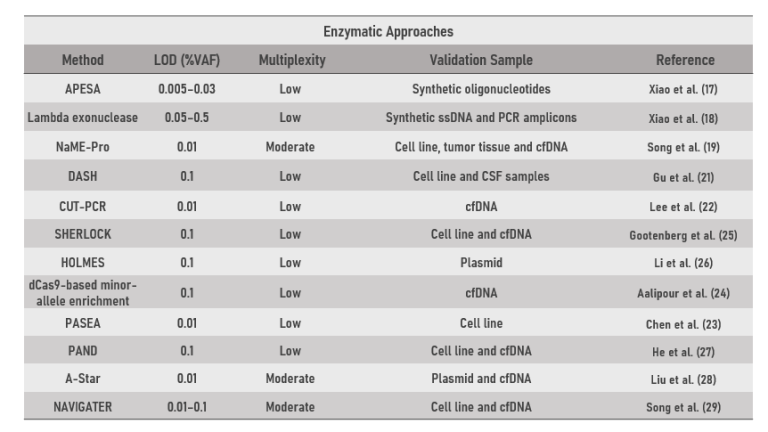
表1 Enzymatic类低频突变特异性富集技术(Multiplexity:Low 1-5/Moderate 100/High 1000 Targets)
基于脱嘌呤/脱嘧啶探针的核酸内切酶IV(Endonuclease IV)信号放大系统(APESA)采用双标记荧光探针,该荧光探针含有与突变模板序列完全匹配的脱嘌呤/脱嘧啶探针(AP探针)[2]。核酸内切酶IV可以识别双链DNA(DSD)中的AP位点,对单链DNA不敏感。因此,当AP探针以最佳Tm与突变链杂交时,核酸内切酶识别并切割探针上的AP位点。随后,AP探针的猝灭基团与荧光基团分离,导致强荧光激发。同时,模板靶序列保持完整,在多次循环后放大荧光信号(图1上)。有趣的是,PCR Blocker(阻断引物结合类)与APESA的信号放大系统结合得到了一种用于富集已知点突变的超灵敏方法(图1下)。通过将PCR Blocker与APESA结合,该技术对C>G和C>A突变的LOD提高到0.005和0.03%VAF。
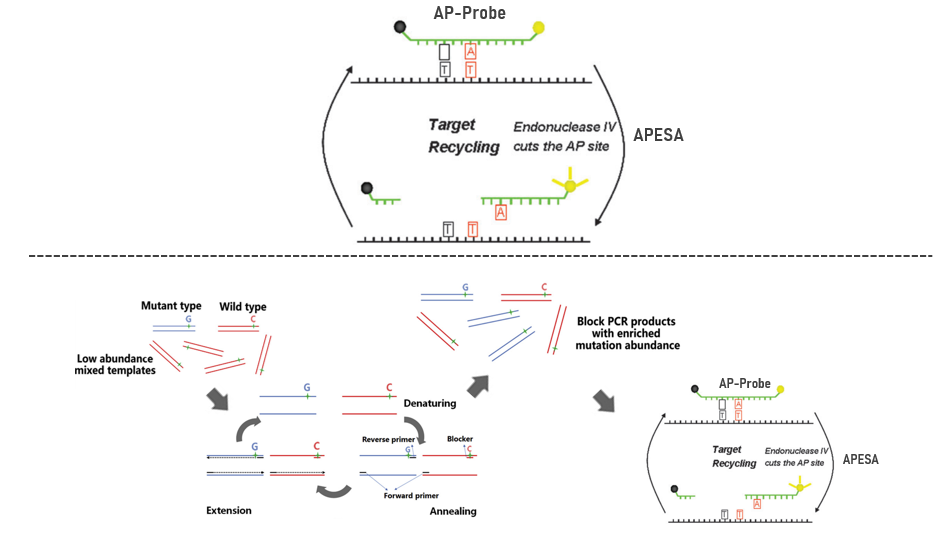
图1 APESA技术原理示意图以及PCR Blocker与APESA的结合技术 [2、3]
在另一种酶学方法中,NaME-PrO技术(nuclease-assisted minor-allele enrichment using probe overlap)采用了一种热稳定的双链特异性核酸酶(DSN, duplex specific nuclease),DSN可以降解完全匹配的双链DNA,而且不具有序列依赖性[4]。为此,针对每个靶点设计一对具有10至15 bp重叠区的wtDNA匹配探针,分别结合正义和反义WT链(图4)。需要富集的突变位点位于探针的重叠部分内,该区域内的任何错配都会导致探针/靶序列的杂交不稳定,从而抑制DSN活性,保留突变DNA模板。NaME PrO技术能够在扩增前在基因组DNA水平上直接对wtDNA进行降解,从而克服PCR过程中聚合酶错误富集wtDNA的潜在问题。该方法为低频突变的多重富集提供了解决方案,可以在不改变其既定工作流程的前提下与下游检测技术结合[5]。

图2 NaME-PrO技术原理示意图以及其对cfDNA进行多重靶点检测结果[4]
另一类酶学法突变富集方法使用中度选择性的CRISPR/Cas系统(clustered regularly interspaced short palindromic repeats/ CRISPR-associated protein)。当CRISPR/Cas核酸酶与引导RNA(gRNAs)偶联,gRNA/靶杂交完全匹配时,可以产生DNA双链切割。CRISPR/Cas核酸内切酶活性需要靶区的3′NGG PAM基序(Protospacer Adjacent Motif)。与NaME-PrO方法一样,CRISPR-Cas9复合物以及针对WT等位基因的gRNA文库能够基本上消除扩增子Pool或二代测序(NGS)文库中的不需要的序列,可以在不增加额外成本的前提下提高突变位点的测序产出,比如DASH(Depletion of Abundant Sequences by Hybridization)和CUT-PCR(CRISPR-mediated, Ultrasensitive detection of Target DNA-PCR)技术 [6、7],DASH和CUT-PCR技术的LOD分别为0.1%和0.01%VAF。

图3 DASH技术原理示意图(左)、运用DASH技术特异降解线粒体核糖体RNA(中)和利用DASH技术特异性富集KRAS G12D突变[6]
该技术的另一种变体是PASEA技术(Programmable enzyme-Assisted Selective Exponential Amplification),将Cas9介导的WT等位基因切割与等温扩增(RPA,Recombinase Polymerase Amplification)结合起来,从而使低频突变等位基因富集达800倍[8]。然而,PASEA要求突变存在于PAM位点才能开展,这大大限制了其应用。与之相反,基于dCas9的突变等位基因富集技术靶向突变序列而不是野生型序列,它包括一种具有His-Tag的无活性重组Cas9与靶向感兴趣突变的gRNA形成的复合物[9]。这项技术使用与抗His抗体结合的磁珠,从而通过免疫法分离提取与靶向突变结合Cas9复合物,LOD可达0.1%VAF。尽管这项技术提供了突变富集,但免疫法捕获和洗涤步骤增加了动手操作时间,并可能导致样品损失和非线性富集。
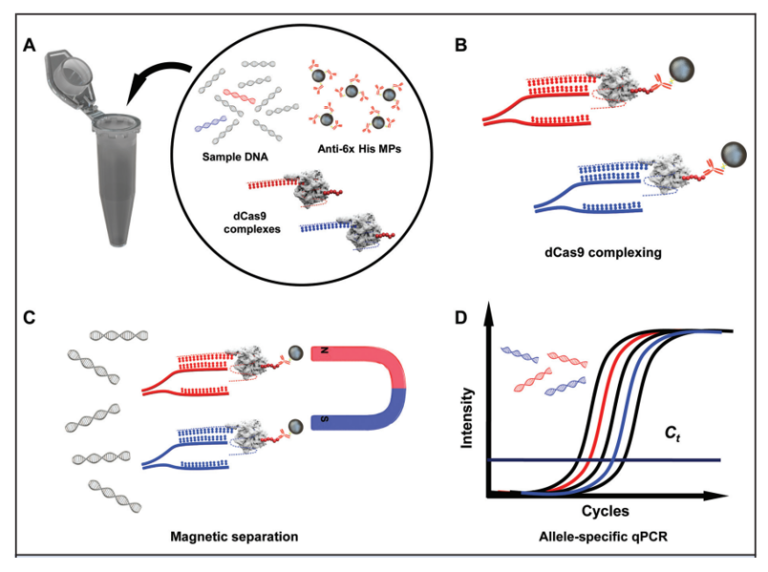
图4 dCas9-based minor-allele enrichment技术原理示意图[9]
除Cas9外,CRISPR效应蛋白Cas13a和Cas12a还被分别用于两种不同的低频突变检测,即SHERLOCK(Specific High Sensitivity Enzymatic Reporter Unlocking; [10])和OLMES(One-Hour Low-Cost Multipurpose highly Efficient System; [11])。这两种方法都基于Cas13a和Cas12a核酸内切酶的一个被称为“collateral effect”的独特活性,一旦效应蛋白/gRNA识别靶序列,Cas13a和Cas12a将激活“无差别攻击”模式,结合并切割环境中的非靶标序列,其不同点在于,SHERLOCK(Cas13a)识别RNA底物,反式切割RNA报告探针,HOLMES方法(Cas12a)识别双链DNA底物,反式切割单链DNA(ssDNA)报告探针(图5),相较Cas13a,Cas12a针对DNA样品时无需体外转录步骤。SHERLOCK和HOLMES技术可以实现0.1%VAF的LOD。
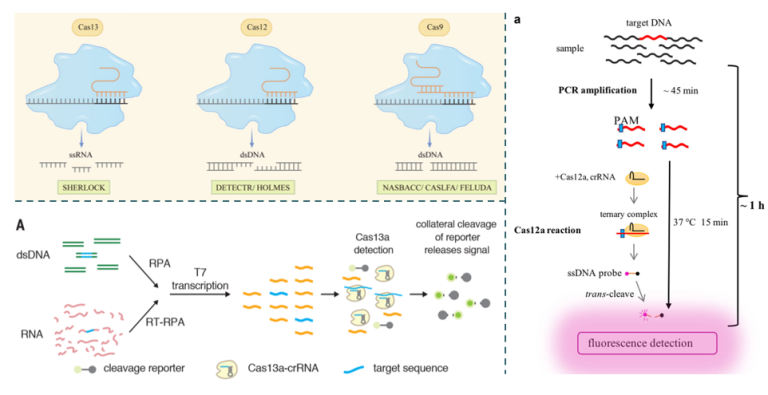
图5 CRISPR/Cas13和CRISPR/Cas12技术原理示意图[10-12]
Cas内切酶的应用受到对靶序列上特定序列基序(比如PAM序列)的严格要求的限制, Argonaute(Ago)内切酶家族没有切割基序要求,而且只需要短DNA引导(gDNA),因此这方面的限制更小。PAND(PfAgo-mediated Nucleic acid Detection,[13])系统使用PfAgo(Pyrococcus furiosus)和3个gDNA切割DNA双链,产生一个16nt的ssDNA,如果该ssDNA含有目标突变,则可以作为新的gDNA(ngDNA,new gDNA),并触发第二轮切割,即ngDNA-PfAgo复合物引导切割事先合成的与突变ngDNA完全匹配的荧光报告探针(图6)。因此,可以通过荧光检测低至0.1%VAF的突变。

图6 PAND技术原理示意图[13]
与之类似的是NAVIGATER技术(Nucleic Acid enrichment Via DNA-Guided Argonaute enzyme from Thermus thermophilus (TtAgo)),TtAgo能够以单碱基的精度特异性地切割互补DNA和RNA,并且对15/16nt长度的引导DNA特异性最佳, NAVIGATER采用TtAgo核酸内切酶对wtDNA进行切割(图7),能够与qPCR、数字PCR和测序平台结合,可以检测0.01% VAF(+XNA-PCR)的突变[14]。

图7 NAVIGATER技术原理示意图[14]
A-Star方法仍然采用PfAgo核酸内切酶,但方式不同[15]。PfAgo具有较好的热稳定性(图8左),该技术在PCR变性步骤中使用PfAgo切割wtDNA的正义和反义链,有效地减少了wtDNA模板(图8右),可以富集低频突变75倍或更多,LOD达到0.01%VAF。该方法在“One-Tube”中完成,是一种多重靶标、灵敏且易于实施的突变特异性富集技术。
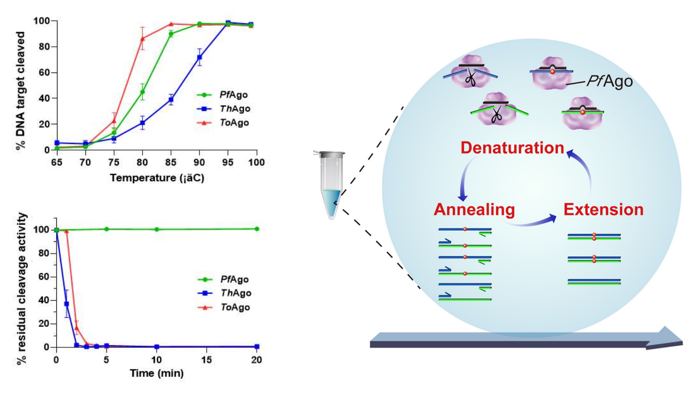
图8 A-Star技术原理示意图以及PfAgo酶活的热稳定性测试[15]
综上所述,酶学法主要是将不同特点的核酸内切酶作为工具,包括1)Endonuclease IV能够特异性识别脱嘌呤/脱嘧啶双链;2)DSN(Duplex Specific Nuclease)则是一种热稳定的双链特异性核酸酶;3)Cas内切酶对靶标序列特征依赖性(PAM等);4)Ago内切酶家族没有切割序列的特征要求;利用它们的特征或直接靶向变异位点序列进行特异性检测,或降解野生型序列间接提高突变序列丰度,从而实现突变的特异性检测(表2)。
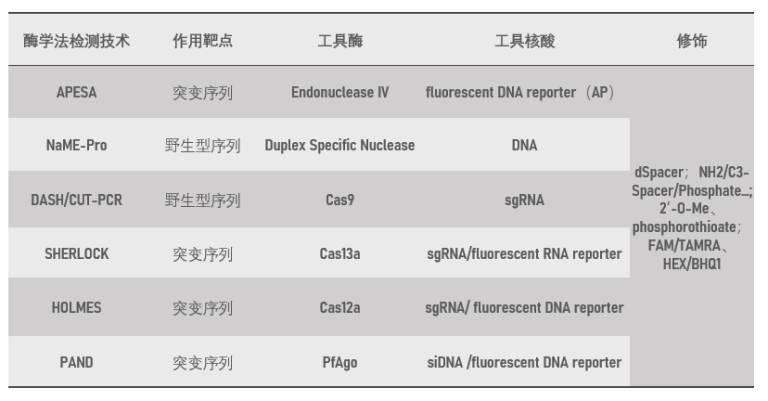
表2 酶学法低频突变特异性富集技术原理特点
伯科生物研发团队自主开发的SynStar21多功能Oligo合成仪,硬件和软件系统可以满足分子检测、核酸药物开发等不同应用方向的DNA和RNA序列合成需求。在酶学法低频突变特异性富集检测应用中,SynStar21多功能Oligo合成仪可以提供DNA和RNA序列的高品质合成,包括DNA/RNA荧光报告探针、引导DNA(siDNA)和引导RNA(转录前DNA或直接RNA合成)等核酸合成。






